The Virtues Of Crossed Connections
Orientation and Object Fixation in Flies
McCulloch-Pitts Neurons and Real Neurons
Shapes: The Morphemes of Visual Perception
An Inborn Category Of Acoustic Form Perception
Structure Of The Cerebral Cortex
Cell Assemblies: Embodiments of Ideas
Threshold Control and the Pump of Thoughts
Braitenberg has presented us with 14 chapters of whimsical thought experiments. At the end of his book, he spends some time making the point that the characteristics that he explored in his vehicles were inspired by a few facts about animal brains, and as a result "their behavior will then seem less gratuitous than it may have seemed up to this point." Let's consider some of these ideas.
This section pertains to Vehicle 2, Vehicle 3, and to Vehicle 4.
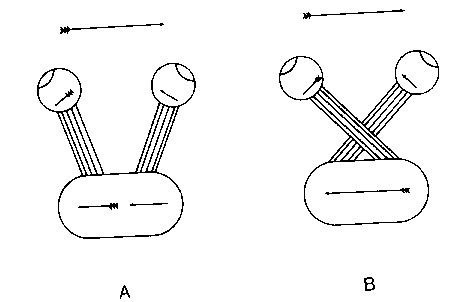
Why does the vertebrate brain have a crossed representation of the world? A variety of explanations have been proposed for this phenomenon. One is that it provides mechanical stability of the brain, because of "strong weaving". Another is that it is the result of the transition from a primitive spherical brain to a brain that is only symmetrical bilaterally. Both of these accounts have their problems.
The best known account was provided by Ramon y Cajal, who argued that crossing corrects the image inversion that occurs because of the geometry of the eye's optics. Specifically, without crossing there would be a midline discontinuity in the mapping of the visual field onto a cortical surface. Such a discontinuity is eliminated by the crossing fibers.
Braitenberg notes that there are several possible objections to this argument. First, crossing is sufficient but not necessary for this correction; in Figure 24 Braitenberg provides several alternatives that could serve the same purpose. Second, the argument presupposes that there is an advantage in not having the midline discontinuity. However, this need not be the case.
In summary, this is still an open topic. I myself have asked neuroscientists about it, and they have not provided any answers. (Question: Has Braitenberg raised alternative answers in describing his vehicles?) They have noted, though, that it is an excellent question!
This section continues Braitenberg's discussion of the reconstruction of continuity in visual representation, and is geared towards Vehicle 8.
Braitenberg has himself provided evidence that the Cajal principle is correct from studying the visual system of the fly. "There the complicated weave of the fibers leading from the compound eye to the brain exactly compensates for the disruption of the image produced by each of the lenses projecting small inverted portions of the visual field onto the array of the light sensitive elements."
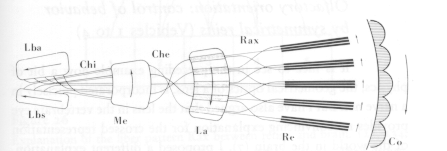
The compound eye of the fly is composed of about 3000 ommatidia. Each has its own optics, and contains 8 light sensitive elements (the rhabdomeres, which are all specialized parts of a single cell, the retinula. The stimulation on the retinula forms a very regular pattern, which consists of 7 lines of sight that intersect a distal plane in a pattern; the pattern of the retinula is this distal pattern rotated 180 degrees. The retinula then projects onto the lamina ganglionaris. There are two properties of this projection that are of interest. First, the nerve bundles involved are literally twisted a full 180 degrees, correcting for the inversion of the retinular pattern. Second, "all the elements that look at the same point of the visual field send their axon into the same compartment of the ganglion", ensuring a continuous projection! There is evidence suggesting that this pattern of connectivity is also innate.
This section pertains to Vehicle 1, Vehicle 2, Vehicle 3, and to Vehicle 4.
Braitenberg suggests that while, from the fly eye example, it is nice to see "how a bit of physics, the geometrical optics of a lens, is incorporated precisely in a nerve net", it is also the case that no such explanation is available for the vertebrate eye. Braitenberg uses olfaction to construct his own explanation.
In the vertebrate brain, the olfactory tracts are not crossed. However, motor outputs are crossed "which means that a certain smell has a stronger effect on the motor system on the side opposite the nostril it hits first." This was reflected in vehicles 2b and 3b. Braitenberg speculates that our vertebrate ancestors may have depended upon olfactory/motor relationships, because of following chemical gradients in the water in which they swam.
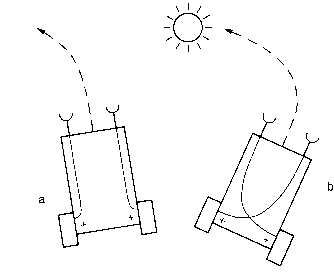
Braitenberg goes on to speculate about this relationship. If these ancestors used fins to propel themselves, then they would be like vehicle 2, where thrust on one side would make the animal turn the other way. In contrast, if they swam like fish via body bends, the reverse would be true.
In short, Braitenberg would like to argue that crossed (or, in some circumstances like olfaction, uncrossed) sensorimotor connections may indeed have been motivated for exactly those reasons that they were incorporated into his early vehicles -- to make the organism move in an appropriate direction when its sensory organs were simulated. Braitenberg goes on to suggest that there does exist some biological evidence (e.g., work on bacteria) consistent with this argument!
This set of notes relates to Vehicle 4, with the key point of reference being the nonlinear function that relates input to output.
Do such relationships exist in neurophysiology? Braitenberg points out that they are indeed quite common, and that when other factors are also taken into account (e.g., changing properties of detectors as a function of position in sensory field) very complicated behaviors emerge. "Or, the other way around, it is sometimes possible to explain astonishingly complex behavior, such as that of a fly navigating through a room and landing on a hanging lamp, by invoking nothing but a set of almost identical, rather simple movement detectors whose output, weighted for position, converges on a few motoneurons."
Braitenberg now moves to additional work on the fly, owing a lot to Reichardt, to elaborate this point. He points out that for flies, detected movement is a key source of information that is crucial to locomotion -- many of a fly's behaviors should be governed by optic flow fields that are detected via the compound eye. But how?
For instance, it is known that flies navigate towards isolated objects that are distinct against a homogenous background. "Generally, perceived motion in the visual field makes the fly turn in the direction of the motion. The effect of the perceived motion may be different for different directions, however. [...] It seems that the difference between the effects of forward and backward motion varies in a systematic way over the visual field. Thus for every pattern in the visual field there will be a net turning tendency." In short, the nonlinearities in the system convert the initial velocity of the fly, and the image details of a room, into the complicated trajectory that the fly takes before it lands. Think again, of Simon's ant!
(NB:One paper that I like a lot, related to this theme, but which people rarely cite, is Dana Ballard's discussion of cortical computation in:
Ballard, D.H. (1986). Cortical connections and parallel processing: structure
and function. Behavioral and brain sciences, 9,, 67-120.)
This section pertains to Vehicle 5, which is identified as an embodiment of the kind of system originally described by McCulloch and Pitts in their classic 1943 paper.
"In their famous paper McCulloch and Pitts stylized the functional relations of neurons connected by synapses as the fundamental operations of the calculus of propositions: conjunction, disjunction, and negation. [...] Is it an accident, then, that conjunction, disjunction, and negation were first defined by the philosophers and then rediscovered as fundamental properties of neurons and synapses in the spinal cord? Or is the nervous system really constructed out of these operations, with the consequence that the philosophers can only discover in their own thinking the laws that make their brains tick?"
Braitenberg admits that it would be practical to speak of neurons and threshold devices as being the same kind of thing, but he doesn't because he views a threshold device as an instance of simplified biology. "Real neurons have properties that go far beyond the simple threshold devices we used as building blocks for our vehicles' brains." In particular, the temporal dimension is very messy in real brains. "The exact point in time at which an action potential arises in a neuron depends not only on the time at which the excitation reaches the neuron but also on the intensity of excitation." This is why putative logic circuit properties of brains are so uncertain. "Whether two spikes will meet or not at a certain synaptic junction, and hence whether the logical operation performed by that junction will occur, may depend on just these unwanted delays."
In the brain, the primary signal is often a burst of several action potentials. Furthermore, the frequency of bursts varies with stimulus intensity -- an analog property. As a result, "there is an analogue principle involved in this which is quite foreign to the digital operation of the Mcculloch-Pitts nerve net, or to the automation of automata theory."
Here Braitenberg reflects back on Vehicle 6. He admits that the selection game that he described in that chapter hushes up most of the complexity of Darwinian evolution. Basically, with these end notes Braitenberg is recommending that psychologists might have something to gain by reading about this theory in order to gain a better appreciation of its complexity.
This section is pertinent to Vehicle 7, Vehicle 10, Vehicle 11, Vehicle 12, Vehicle 13, and to Vehicle 14.
He admits that the Mnemotrix and Ergotrix wires are flights of fancy (surprise, surprise), but argues for their important functional role. He notes that these vehicles depend heavily upon Hebb's original ideas. Finally, he suggests that his vehicles (and related models) "were all created under the impression that `association' is the most important principle by which information about the environment is incorporated into the brain."
In previous notes, I hinted that Braitenberg's reliance on associationism was a weak link in his chain. He admits that association may not be a powerful enough theory to account for the effects that must be explained in cognitive psychology, and that new information about the complexity of neuron function also suggests that reliance on pure association is too simplistic. Sutton and Barto are key references for this claim.
He then goes on to ask whether there is any direct physiological evidence for the kind of learning that was incorporated into these machines. He argues yes, and gives some evidence, but all of this was clearly written prior to current advances in our understanding of LTP and LTI, and the underpinning of NMDA receptor mechanisms. If anything, stronger evidence -- but more complicated evidence -- exists for his view of learning.
This section is pertinent to Vehicle 7, and to Vehicle 11.
His main point is that it almost goes without saying that learning involves "some anatomical rerouting, that is, changes in the synapses of the network." He wants to stress this point because it has not always been accepted. "It is no longer fashionable, luckily, to imagine that the information of complex experiences resides in individual molecules of the brain." Indeed, from reading his comments on this, I get the sense that such molecular theories of learning are pet peeves of Prof. B.
I think that what concerns Braitenberg is that his view -- learning = synaptic modification -- is not as shored up as he would like. He spends a couple of pages to make the point that there are deep problems with isolating the physical nature of the memory trace. Really, one gets a sense that what B. desires is a physical account of the specific content of memories.
My own personal slant on this issue is this: most connectionist systems model learning as a kind of synaptic change. However, it is almost assumed a priori by connectionist researchers that this kind of change is distributed in a network in a fashion that identifying the physical properties associated with a particular memory trace (e.g., look at the physical properties, and then infer the content) is an impossible task.
These notes pertain to Vehicle 8 and Vehicle 9, which Braitenberg acknowledges "have not sprung merely from creative fancy." Indeed, they reflect "the main theme of brain research for the past hundred years: the representation of external spaces in the spatial coordinates of the nervous system."
Braitenberg raises one really neat idea here. He notes that coordinate systems do not necessarily play a role in some computations, such as language or abstract thinking. Where does this non-geometrical computation take place?
Braitenberg is cautious about arguing that the creation of topological representations is necessarily due to their importance for geometric computations. For example, he points out that such representations may be an artifact of embryology, where as fibre bundles grow neighborhood relations are merely preserved (ie., preserved for no reason other than the fact that there are neighboring fibers). But Braitenberg strongly feels that this is not the whole story, that biological maps emerge because of the special kinds of computation that they can mediate. (One gets a particular sense of this when specific topologies, like those discussed previously for the fly, look very bizarre until taken into the context of the optical system that delivers the proximal stimulus.)
 Braitenberg's 4D space
Braitenberg's 4D space
Another interesting point, about Braitenberg's drawing of a four-dimensional space in a network -- "networks may be symmetric in any number of dimensions and still be housed comfortably in the 3-dimensional space surrounding us, or even in the 2-dimensional space of a drawing. This was just intended as a warning to neuroanatomists who cannot abstract from what they see." In short, Braitenberg would not be surprised if someone was able to describe a tissue as a very high dimensional topographic map, though no one has done this yet.
This section is pertinent to Vehicle 9.
Braitenberg dedicates vehicle 9 to the memory of Gestalt psychology, which B. defines as "the study of the concept of shape." In spite of his admiration for the Gestaltists, Braitenberg notes that "Gestalt psychologists were not so successful, however, in their attempts at relating their discoveries to functional principles of brain physiology." Even today, Braitenberg laments the fact that we are just beginning to understand "the codewords the brain uses in categorizing shapes."
One example that Braitenberg focusses on is "clustering", or "cluster detection", where a number of independent stimuli are grouped into a unitary whole. Braitenberg argues that a neuroanatomical analog of cluster detection is found in the dendritic trees of purkinje cells, where cover an area an order of magnitude larger than the distance between cell bodies with several thousand synapses in the dendrites. "Thus each responds to the activation of a cloud of synapses centered around it, and the clouds of synapses belonging to neighboring neurons overlap generously."
From this observation, Braitenberg considers scale of resolution, arguing that what is detected as a single cloud by large scale neurons can be detected as a set of individual points by smaller neurons -- a size of filter effect. "We are able to switch rapidly from one set of filters to another, making available to the form-perceiving mechanism different bands of the space-frequency spectrum." (NB:This makes me think of Marr's view of processing from the raw to full primal sketch, which depends a very great deal upon low frequency filters constraining the activity of higher frequency filters.)
Braitenberg discusses a neural network for trajectory tracking, and then raises the possibility that it might implement a natural constraint to distinguish real objects from random noise etc., "for the most common thing that can be said about physical objects is that they move at reasonable speeds without breaks in their trajectories." Personally, I am very skeptical that this notion could serve as a natural constraint.
All in all, we see that Braitenberg envisages the use of computer simulation to create a synthetic psychobiology. "We can no longer fondle our ideas about the brain with the secure feeling that their falsification is beyond technical feasiblity. Most ideas can be translated into computer programs and are thus easily put to the experimental test."
Nevertheless, much of how the brain does perception is still a mystery.
This section is pertinent to Vehicle 8 and to Vehicle 9.
There is lots of evidence of inborn perceptual categories, and their relationship to neuroscience. Braitenberg discusses one very briefly, the notion that a melody played in a different key sounds pretty much the same to us. Braitenberg argues that what underlies this is the notion that in the brain, sound frequencies are represented on a logarithmic scale. "The resulting translational symmetry for tone patterns characterized by constant frequency ratios is one of the basic facts of music."
This section is pertinent to Vehicle 11.
For such more sophisticated vehicles, Braitenberg argues that it is much more difficult to justify their architecture by pointing to known properties of the brain. Here, Braitenberg is reduced to playing a kind of numbers game. By considering the probable number of connections involving interneurons, and considering the probably number involved in direct contact with the world, "we may infer that the internal, cortico-cortical connections of the cortex are at least 10 times, perhaps a 100 times more powerful than the connections of the cortex with the external world."
So what does all of this mean? Well, it appears to be a characteristic cortical fact. And from this, plus a detailed description of pyramidial cells, Braitenberg seems to say that this might give enough associative horsepower to mediate higher-order functions.
This section pertains to Vehicle 7 and to Vehicle 10.
Braitenberg suggests that it is fashionable to think of cell assemblies as the carriers of individual items of meaning in the brain -- as representational primitives. However, he also notes that there is no logical reason for this view. Instead, the reasons appear to be empirical:
"First, many years of recording responses of single neurons to sensory stimuli have shown that no very complicated or very unique input is needed to activate a neuron." Simple features are the norm. Because they can't be very meaningful, this suggests that sets of neurons -- each sensitive to different simple features -- are required for complex representation. (NB: This reminds of the work that I did 10 years ago on analyzing alphabetic confusion matrices; work that nicely fit with the notion that perception is the accumulation of simple features.)
Second, observations of the plasticity of synapses is consistent with the notion that Hebb learning occurs, coupled with Braitenberg's suggestion that most interneuron connections are excitatory in nature. (NB: I have to dig this up, but I am sure that some circa 1960s computer simulations on Hebb's model indicated that inhibition was required for anything to work at all.)
Third, Braitenberg would have us think that the sheer size/complexity of cortical wetware is nicely consistent with the reality of cell assemblies.
This section is pertinent to Vehicle 12, Vehicle 13, and to Vehicle 14.
Threshold control winds up being a critical component in Braitenberg's later vehicles, which is hardly surprising given their associationistic nature. Is there any evidence for this kind of regulation in the brain? Braitenberg notes that recent evidence is consistent with the notion that the caudate nucleus might detect overall cortical activity, and thus be able to send signals to modify thresholds.